
Swimming: the act or art of sustaining and propelling the body in water.1
On our blue planet, many life forms were once faced with the same question: how do I move through that aqueous medium towards that prey/shelter/sexual partner over there? What I want to discuss here is how many possible answers there are to this question.
First, let’s look at us, humans. On the top of my head, I can name at least three different swimming styles: breaststroke, butterfly, front crawl. Two things come to mind here:
- First, there are obviously several ways to swim, if you can find at least three different ones in a single tetrapod species. Throw in some mollusks and osteichthyans and for sure we should find hundreds more.
- Second, some ways are more efficient than others. There’s a reason every single athlete at the Olympic event of “freestyle swimming” will swim front crawl: it’s just faster.
So, in the great freestyle swimming competition of evolution, is there a universal solution? May I suggest we take a look at a video by Code BH, a programmer who simulated the evolution of swimming in aquatic creatures.
I think this video is great. It’s fun, it’s well explained, and it’s quite interesting scientifically. It’s in French, and I didn’t find English subtitles, so I will sum it up here for potential non-francophone readers, and to highlight some points.
Jellyfish versus snake
The problem of swimming appears to be a simple one: pushing against a fluid that exerts an opposite force when pushed against. Let’s call this property the viscosity of water2. The programmer applies an algorithm that selects for swimming efficiency on 2D-articulated, randomly generated creatures. In this program, the creatures that swim the fastest while spending the least energy go on to the next generation and reproduce (with some mutations).
Early on in the simulation (meaning: not much selection yet), the most efficient swimmers are “jellyfish”. They thrive because their shape is simple, just 2 segments joined by one articulation, and therefore the chance they randomly move their articulations in a swimming-compatible manner is higher than for more complex creatures.
Not all “jellyfish-like” creatures are good swimmers: some move their segments (from here on: “arms”) in a way that keeps them in the exact same place. The key thing with the “jellyfish strategy” is that (1) the 2 arms need to close fast to expel water and propel the creature forward, then (2) stay closed to keep the creature in a hydrodynamic posture and finally (3) slowly reopen the arms, before quickly closing them again, and so on. What does the trick here is inertia: the movement generated at step (1) continues during step (2).

But, as the simulation goes on and selection acts, the jellyfish are quickly replaced by more efficient competitors: the “snakes”. The “snake strategy”, that requires a creature with at least 3 segments, goes something like this: alternate movements of the articulations up and down to create an undulation that propels the creature perpendicularly to the movements (as opposed to longitudinally for the jellyfish).
Basically, the snake strategy eliminates the glaring flaw of the jellyfish strategy: its step n°(3). When it reopens its arms, the jellyfish inevitably pushes water forward, therefore applying a force that drives it backward (because viscosity). While the jellyfish can optimise its movement with perfect timing, it cannot escape this step that costs energy to generate a force in the wrong direction. In the snake strategy, every movement contributes to the continuation of the undulation, and therefore every bit of energy is used to propel the organism forward.

With simple, randomly generated creatures, this video shows us that natural selection tends to favor one way of swimming that is the most efficient, based on undulations of the body. We can see variations of this strategy in many sea organisms, from dolphins to tuna to sharks to… well, sea snakes, obviously.
The magical number 20
But of course, jellyfish exist in reality, because natural selection does not select exclusively on swimming efficiency (if that was the case, we would live in a world exclusively populated by Léon Marchands and Katie Ledeckys). Suboptimal ways of swimming can persist if they appear in bodies that otherwise fit an ecological niche.
Some animals like rays or cuttlefish swim thanks to ribbon-like fins that undulate along their body. This is not the fastest way of swimming, but it works well enough for organisms with a rather flat body. In fact, this type of locomotion has evolved independently in at least 4 very distant families of animals.

This is a beautiful illustration of the process called convergent evolution : when faced with the same constraints, different organisms can develop similar solutions because natural selection will favour similar outcomes. How similar you ask? Well…
Here’s a very nice article by Rahul Bale and his colleagues in PLoS Biology: “Convergent Evolution of Mechanically Optimal Locomotion in Aquatic Invertebrates and Vertebrates” (2015). They measured a bunch of biophysical parameters in videos of organisms that swim with elongated fins (technical term: “median/paired fin” swimming). One striking finding: all these animals, having absolutely nothing to do with each other phylogenetically, undulate their fins at the same wavelength. More precisely, the ratio of the wavelength of an undulation to its mean amplitude (called the specific wavelength, SW) is consistently found to be around 20.
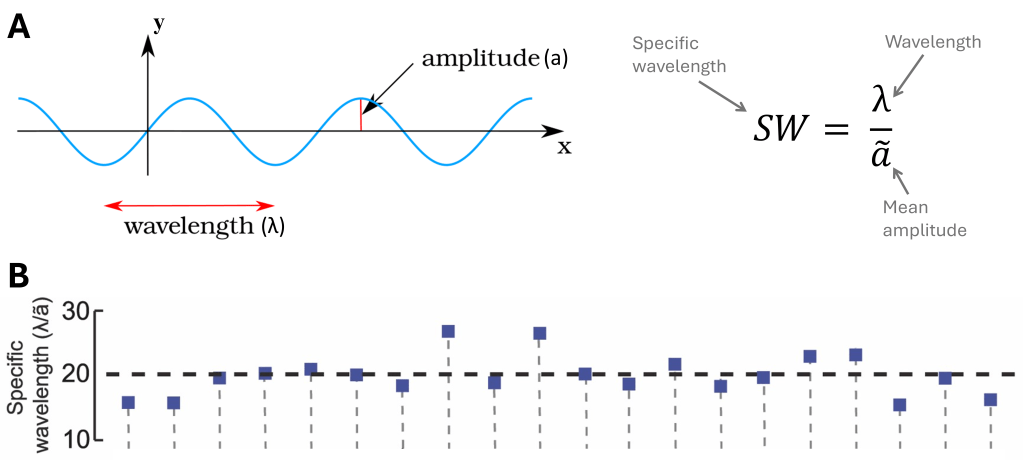
Why 20 ? Bale and team ran a series of tests with a computer simulation of a robotic knifefish3 they built. The results are crystal clear: in a fluid with the density and viscosity of water, whatever the fin height or length, frequency of undulations or angular excursion4, the maximum propulsive force is always reached for a SW value of 20.
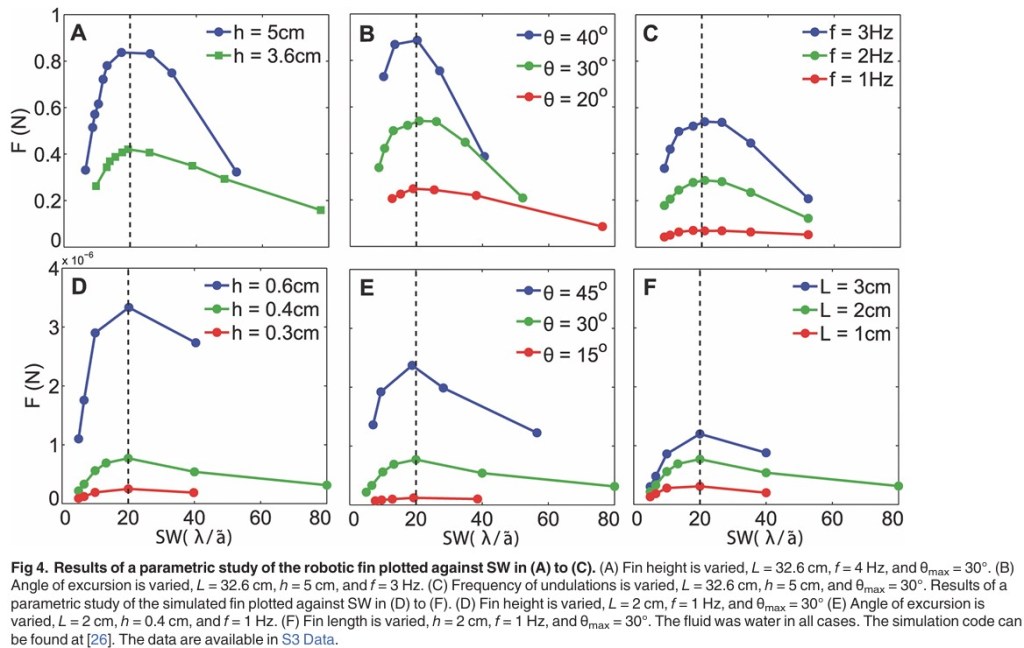
Even within the suboptimal style of “median/paired fin” swimming, natural selection has forced aquatic life forms to adopt a specific wavelength of undulation that optimises the swimming motion. This is an example of how the laws of physics may channel evolution towards a certain path, regardless of biological diversity. For a last and even more drastic example of this, let us turn, once again, to microbial life.
The impossibility of a microscopic scallop
The third resource I’ll examine in this post is a kinda weird paper from 1977 by Edward M. Purcell. Weird, because it is the retranscription from a talk given by the author at a physics seminar, with figures reproducing the transparencies used during the talk. It’s bloody awesome and fun, and I wish more papers were like this5.
This talk/paper is titled Life at low Reynolds number. The Reynolds number (R) is a physical quantity related to a system in a fluid, that represents the ratio of inertial forces to viscous forces in said system. Here’s a small explanatory figure:
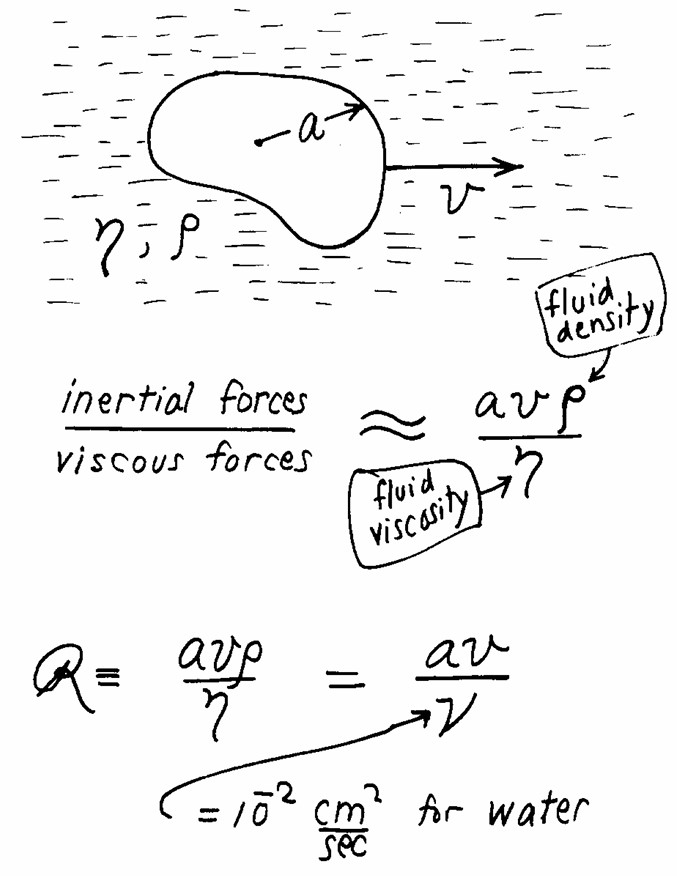
For a system of dimension a, moving at speed v in a fluid of density rho and dynamic viscosity eta, the Reynolds number is the ratio [(a.v.rho)/eta]. The consequence of this formula is that 2 systems that differ in size and speed, moving in the same fluid, will have different Reynolds numbers. Here’s a nice illustration of this:
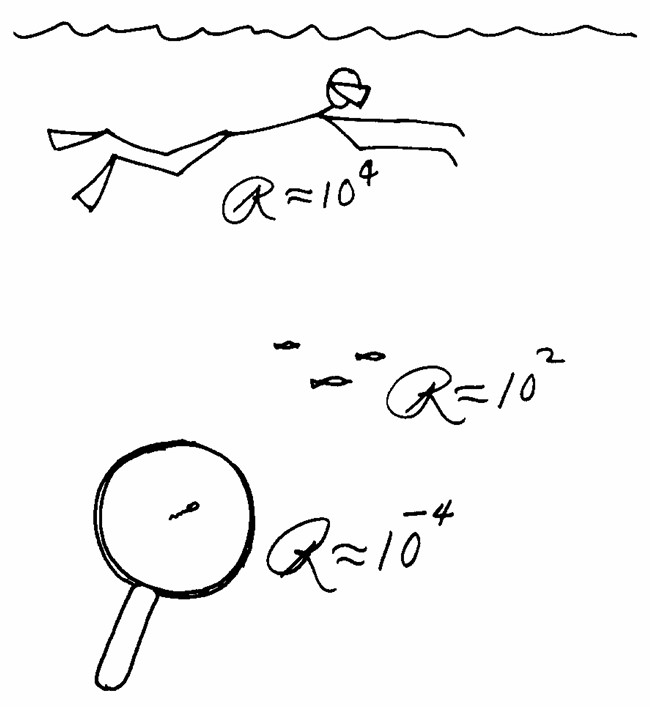
A whale and a marine bacterium live in the same medium, seawater, yet the ways they experience moving in seawater are fundamentally different. This is because a 1 µm-long bacterium and a 15 m-long whale, as physical systems, have respective Reynolds numbers separated by 14 orders of magnitude. Because R is the ratio of inertial forces to viscous forces, creatures at very low Reynolds numbers experience virtually no inertia. In fact, our 1-µm long flagellated bacterium swimming at 30 µm/s6, if it suddenly stops swimming, will drift for only 0.3 µs, for a distance of 0.1 Ångstrom7. That’s about 1/10th of the diameter of a hydrogen atom.
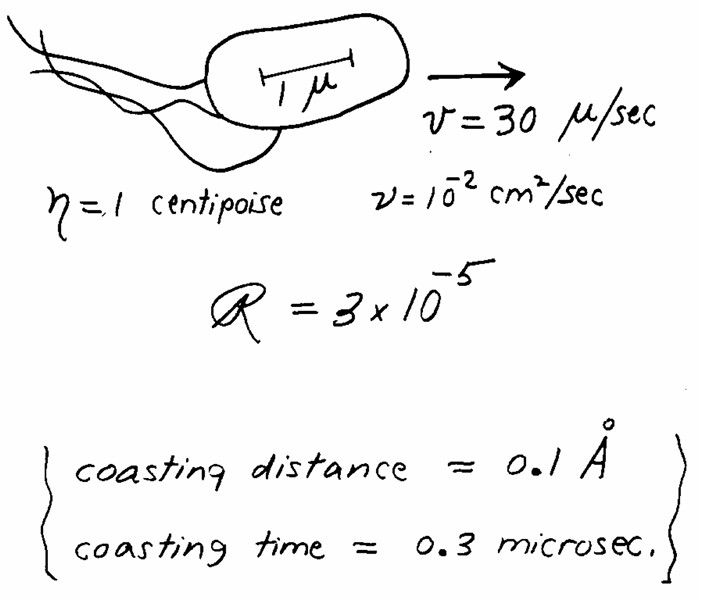
What does it mean to swim in a world with no inertia? The speed you experience at time t will be completely determined by the force you produce at time t, and completely independent of the force produced at time t-1s. To borrow an analogy from Purcell, imagine swimming in a pool of honey. This is a pretty drastic constraint imposed on microbial movement.
One of the major implications is that, at low Reynolds numbers, any movement that depends on returning to an initial state via a reverse motion (example: closing the arms fast then reopening them slowly) is bound to go back to its initial position. As Purcell puts it:
Time, in fact, makes no difference – only configuration. If I change quickly or slowly, the pattern of motion is exactly the same.
[…]
So, if the animal tries to swim by a reciprocal motion, it can’t go anywhere.

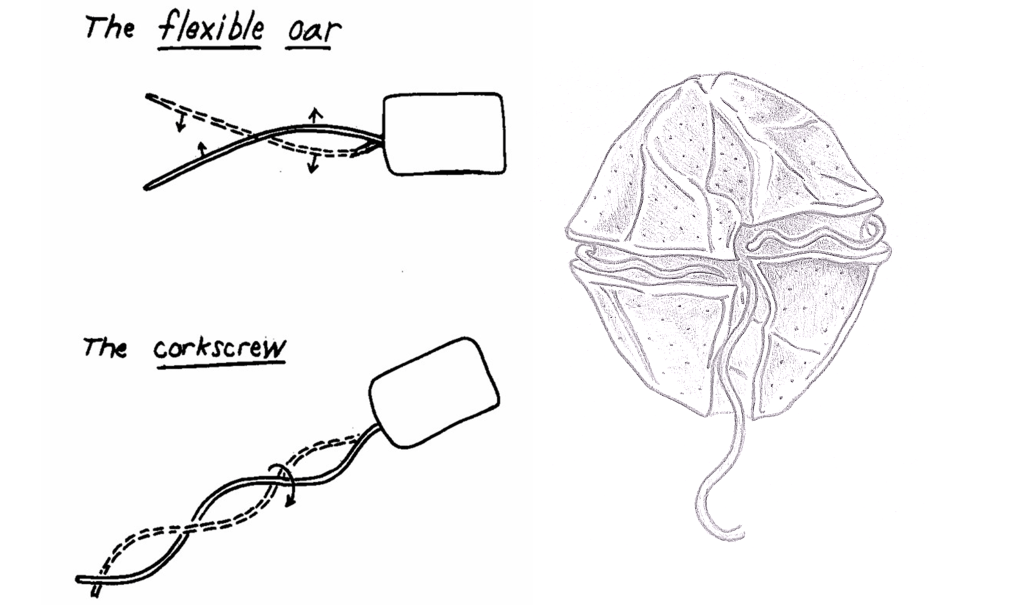
Purcell calls this “the scallop theorem“, because it applies perfectly to the movement of a scallop, that quickly closes the two valves of its shell to propel itself and then reopens them slowly – in other words: the scallop is an adept of the jellyfish strategy. The theorem states that microscopic organisms cannot swim with the jellyfish strategy. A microscopic scallop is impossible.
Therefore, most swimming microbes follow the snake strategy, with the undulating movement of a cellular appendix, usually a flagellum (or more than one)8. Not because this is the optimal swimming strategy in the microbial world, but because it is the only viable one.
Life rebooted
What should we take from this?
First and most importantly, anyone currently searching for microbial scallops should stop immediately.9
Second, let’s quote the first sentence from Bale et al. (2015):
How would life look if it evolved again on Earth, or for that matter, on any other habitable planet?
If life was rebooted from LUCA, or even from the very first cell, on an exoplanet, we might expect that evolution, acting through the randomness of mutations, would give rise to very alien-looking life forms. And at the same time, because the laws of physics are the same everywhere in the universe, we should also see similar structures and strategies emerge, in adaptation to the same fundamental constraints.
There is a famous book by French biologist Jacques Monod titled Chance and Necessity10. The characteristics of living beings evolve by chance, but for them to endure it is necessary that they pass the test of natural selection. Despite the astounding biodiversity on Earth (and, who knows, maybe elsewhere), there is not an infinite number of answers to the question “How can I swim?”. In some situations even, there is only one.
And if you ever happen to get stuck in that pool of honey, well, just keep swimming.
This post examines and uses extracts of the following video and two articles:
- J’ai simulé l’évolution de créatures aquatiques. Video published on the YouTube channel Code BH on March 23, 2023. (https://youtu.be/gVEWaOtEASM?si=Ot0w_n0m1vK8_vRX)
- Bale, Rahul, et al. “Convergent evolution of mechanically optimal locomotion in aquatic invertebrates and vertebrates.” PLoS biology 13.4 (2015): e1002123. (https://doi.org/10.1371/journal.pbio.1002123)
- Purcell, Edward M. “Life at low Reynolds number.” American Journal of Physics 45.1 (1977).(https://www.damtp.cam.ac.uk/user/tong/fluids/lowreynolds.pdf)
I would also like to acknowledge 3 people who indirectly contributed a lot to this post. First, the author/artist behind this beautiful blog post that led me to the discovery of the Bale et al. (2015) article on convergent evolution of swimming. Second, many thanks to Òscar Guadayol who introduced me to the marvels of microbial physics (and to the Purcell paper!) in a very nice seminar he gave during his visit in Brest last spring. Finally, thanks to my flatmate Augustin for showing me the video on simulating the evolution of aquatic creatures.
Any illustration in this blog post identified as “CC BY” is under a Creative Commons Attribution license and can be reused freely as long as the author(s) is (are) properly acknowledged. Other illustrations may be subject to copyright.
- definition in the wiktionary ↩︎
- This is probably a gross simplification, but I’m unrecoverably bad at physics, as will appear more and more clearly as this post goes on. If you’re a physicist and spot a glaring error, please don’t hesitate to correct it by posting a comment! ↩︎
- Between the words “a computer simulation of a robotic knifefish” and the incredible name Pseudobiceros fulgor, I don’t know how this post can get any more sci-fi. ↩︎
- (The angular excursion measures how far the fin can deviate from the body plane. Basically, it’s a measure of the flexibility of the fin.) ↩︎
- Which makes it even worse that this 49-year-old paper is not open access, and on sale on the website of AIP publishing for 40 FUCKING DOLLARS. For God’s sake. Anyway, I’m certainly not encouraging you to go look for it on sci-hub (where you’ll certainly not find it anyway), because this would definitely be a very bad thing to do. Fortunately you won’t have to, because this link to a nice pdf version was generously provided by Jeremy Fox in a recent post on the Dynamic Ecology blog. ↩︎
- Which is an impressive speed if you think about it: it’s moving 30 times it’s body length in one second! ↩︎
- 1 Å = 10-10 m. ↩︎
- I guess the ciliae of ciliates could be described as a multitude of small flagella? Also, I’m excluding the movement of benthic diatoms because they glide on a solid surface (and are therefore not “swimming”). ↩︎
- However, some ciliates (for instance, good ol’ Mesodinium rubrum) seem to move by jumps, which puzzles me a bit. A paper analysing the motion of Mesodinium cells (Fenchel and Hansen, 2006) seems to find that the jump involves some rotation, so maybe this is a kind of superfast corkscrew? ↩︎
- Chance and Necessity: Essay on the Natural Philosophy of Modern Biology, Jacques Monod, (1970). En français : Le Hasard et la Nécessité. ↩︎

Leave a comment